
In Silico Profiling of Cytokine Signatures in Hepatocellular Carcinoma Reveals Elevated CXCL9/10 and Reduced IL-10 Expression
Abstract
Introduction
Hepatocellular carcinoma (HCC) arises within a pro-inflammatory microenvironment where cytokines and chemokines significantly influence tumor development and immune response. This study aimed to identify specific immunoregulatory alterations associated with malignant transformation in HCC by analyzing transcriptomic differences in cytokine and chemokine gene expression between tumor and adjacent non-tumor liver tissues.
Methods
An in silico transcriptomic analysis was conducted using the publicly available RNA-seq dataset GSE124535, which includes 73 paired HCC (tumor, T) and adjacent non-tumor (P) liver samples. Expression levels (FPKM) of 13 cytokine and chemokine genes, IL1B, TNF, IL6, IL10, IL1RN, TGFB1, CCL2, CXCL8, CXCL9, CXCL10, IFNG, IFNA1, and IFNB1, were quantified and transformed using log2(FPKM+1). Statistical significance of differential expression was evaluated using Mann–Whitney U tests and Welch’s t-tests. Principal component analysis (PCA) was applied to Z-score–normalized cytokine expression data.
Results
CXCL10 expression was significantly elevated in HCC samples compared to non-tumor tissue (mean log2[FPKM+1]: 4.35 vs. 3.15; p = 1.3×10−3, Wilcoxon; p = 9.6×10−4, t-test), alongside a moderate increase in CXCL9 (3.30 vs. 2.58; p ≈ 3.0×10−2 and 2.2×10−2). In contrast, IL-10 expression was significantly decreased in tumor tissue (0.31 vs. 0.78; p = 2.9×10−4 and 7.9×10−5). Other cytokines did not exhibit statistically significant differences. PCA revealed partial separation between tumor and non-tumor samples, with PC1 and PC2 explaining 32% and 18% of total variance, respectively.
Discussion
The observed cytokine expression profile in HCC suggests a shift toward enhanced chemotactic signaling, particularly via CXCL9 and CXCL10, coupled with reduced anti-inflammatory regulation through IL-10. This altered cytokine landscape may influence tumor immune evasion and shape the tumor microenvironment, highlighting the importance of these mediators in disease progression.
Conclusion
HCC is associated with a distinctive cytokine signature characterized by the upregulation of chemokines CXCL9 and CXCL10 and downregulation of IL-10. These findings indicate that CXCL9 and CXCL10 may serve as promising diagnostic and prognostic biomarkers, while IL-10 represents a potential therapeutic target for modulating the immune response in HCC.
1. INTRODUCTION
Hepatocellular carcinoma (HCC) is the predominant form of primary liver cancer and a major contributor to cancer-related mortality worldwide, accounting for over 905,000 deaths in 2022 and ranking third in global cancer mortality [1]. The incidence of HCC has been steadily increasing over the past four decades, driven primarily by chronic hepatitis B and C virus infection, aflatoxin exposure, and the rising prevalence of non-alcoholic fatty liver disease (NAFLD) [2]. Despite advances in diagnostic imaging and surgical techniques, the prognosis remains poor, with a five-year survival rate below 20% for advanced-stage disease [3]. Limited efficacy of current systemic therapies and the complex interplay between tumor cells and the liver microenvironment underscore the urgent need to identify novel biomarkers and therapeutic targets.
Inflammation is a hallmark of HCC pathogenesis, as chronic liver injury and regeneration create a pro-tumorigenic microenvironment that promotes genetic mutations, epigenetic alterations, and immune dysregulation [4]. Central to this process are cytokines and chemokines-small secreted proteins that mediate intercellular communication and orchestrate immune cell trafficking, activation, and differentiation. Pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6) activate oncogenic signaling pathways, NF-κB and STAT3, respectively, that drive hepatocyte proliferation, survival, and resistance to apoptosis [5]. Conversely, immunosuppressive cytokines, including interleukin-10 (IL-10) and transforming growth factor-beta (TGF-β), contribute to tumor immune escape by inhibiting effector T cell responses and promoting regulatory T cell and myeloid-derived suppressor cell accumulation [6].
Chemokines, a specialized subset of cytokines, regulate leukocyte recruitment to the liver and are implicated in both anti-tumor immunity and tumor progression [7]. For example, CXCL9 and CXCL10 bind the CXCR3 receptor to attract cytotoxic T lymphocytes and natural killer cells, yet paradoxically elevated levels in the HCC microenvironment have been associated with poor prognosis, possibly due to chronic overexpression leading to immune cell exhaustion or aberrant angiogenesis [8]. Moreover, CCL2 and CXCL8 have been shown to recruit tumor-associated macrophages and neutrophils that support tumor growth, invasion, and metastasis [9]. Understanding the balance and context-dependent roles of these mediators is critical to designing effective immunomodulatory strategies in HCC. Recent evidence suggests that dysregulation of these chemokines, particularly CXCL9 and CXCL10, could be exploited as diagnostic or prognostic biomarkers, providing valuable information for disease staging and patient stratification [10].
In parallel, the immunosuppressive cytokine IL-10 has emerged as a key regulator of tumor–immune interactions, and its modulation is increasingly being explored as a therapeutic strategy. Targeting IL-10 signaling pathways may restore anti-tumor immune activity and enhance the efficacy of immunotherapy in HCC, underscoring the clinical importance of characterizing its expression dynamics.
Transcriptomic profiling has emerged as a powerful tool to dissect the molecular landscape of HCC. Several studies using microarray and RNA-sequencing platforms have identified gene expression signatures associated with tumor grade, vascular invasion, and patient survival [11]. However, comprehensive analysis of cytokine and chemokine expression across matched tumor and adjacent non-tumor tissue remains limited. Publicly available datasets such as GSE124535 offer an opportunity to perform systematic, in silico analyses of cytokine expression profiles, enabling hypothesis generation without the need for additional wet-lab experiments. Combining robust statistical testing with principal component analysis and pathway enrichment can reveal coordinated shifts in immune mediators that characterize the HCC microenvironment.
In this study, we profile thirteen key cytokines and chemokines, IL1B, TNF, IL6, IL10, IL1RN, TGFB1, CCL2, CXCL8, CXCL9, CXCL10, IFNG, IFNA1, and IFNB1, in 73 paired HCC and adjacent non-tumor liver samples from the GSE124535 RNA-seq dataset. We apply log2 transformation and both parametric and nonparametric statistical tests to identify differentially expressed genes, followed by principal component analysis to assess the utility of the cytokine signature in distinguishing tumor from normal tissue. Our analysis uncovers a distinct HCC-associated cytokine landscape characterized by elevated CXCL9/10 and reduced IL-10 expression, suggesting an imbalance between chemotactic signaling and anti-inflammatory regulation. These insights not only deepen our understanding of the immunological dynamics driving HCC but also point toward novel biomarker candidates and immunotherapeutic targets with potential clinical utility.
2. METHODS
2.1. Study Design
This analytical observational study employed a quantitative, in silico approach to evaluate cytokine and chemokine expression signatures in hepatocellular carcinoma (HCC). The research aimed to identify immunoregulatory gene expression alterations associated with malignant transformation in HCC by comparing tumor tissue with adjacent non-tumor liver samples. Publicly available RNA sequencing data from the Gene Expression Omnibus (GEO) repository (accession number GSE124535) were analyzed. The dataset comprised 73 paired tumor (T) and adjacent non-tumor (P) liver samples. Expression values for thirteen cytokine and chemokine genes-IL1B, TNF, IL6, IL10, IL1RN, TGFB1, CCL2, CXCL8, CXCL9, CXCL10, IFNG, IFNA1, and IFNB1-were retrieved in fragments per kilobase of transcript per million mapped reads (FPKM) format and transformed using log2(FPKM+1). Group-wise comparisons were performed using both Mann–Whitney U tests and Welch’s t-tests to identify statistically significant differences in gene expression. Z-score–normalized data were subjected to principal component analysis (PCA) to explore sample clustering and variance in cytokine expression patterns. This non-experimental, retrospective design provides insight into cytokine-driven immune signatures in HCC and supports the identification of candidate immunological biomarkers.
2.2. Search Strategy and Data Selection
A systematic search of publicly available transcriptomic datasets was performed using the NCBI Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/) to identify RNA sequencing studies relevant to hepatocellular carcinoma (HCC). The search strategy incorporated the following keywords and Boolean operators: (“hepatocellular carcinoma” OR “HCC”) AND (“RNA-seq” OR “transcriptome”) AND (“tumor” AND “non-tumor” OR “adjacent tissue”). Filters were applied to restrict results to Homo sapiens, RNA-seq platforms, and datasets that included paired tumor and adjacent non-tumor liver tissue samples. Studies were included if they provided raw or normalized RNA-seq data (e.g., FPKM or TPM), had paired tumor and non-tumor samples from the same HCC patients, contained clearly annotated sample metadata, and were publicly available for reanalysis. Datasets were excluded if they lacked a paired design, used microarray or non-transcriptomic platforms, focused on cell lines, animal models, or organoids, or lacked sufficient tissue annotation or metadata. Based on these criteria, GSE124535 was selected due to its high-quality paired design and adequate sample size (73 patient-matched pairs). This dataset includes RNA-seq profiles of tumor (T) and adjacent non-tumor (P) liver tissues from individuals with HCC. The expression data were downloaded in FPKM format and subsequently preprocessed for downstream quantitative analysis.
2.3. Inclusion and Exclusion Criteria
To ensure transparency in dataset selection, explicit inclusion and exclusion criteria were applied at both the study and sample level. Because this investigation was based on secondary analysis of publicly available RNA-sequencing data, eligibility was determined by evaluating the design, metadata quality, and suitability of candidate datasets retrieved from the NCBI Gene Expression Omnibus (GEO). Only datasets involving human hepatocellular carcinoma (HCC) and containing paired primary tumor tissue alongside matched adjacent non-tumor liver tissue were considered eligible. Studies were required to provide RNA-seq data in raw or normalized formats (FPKM, TPM, or count matrices) that permitted quantitative reanalysis and log2 transformation. Clear and complete sample annotations were necessary to allow unambiguous classification of each specimen as tumor or non-tumor. Datasets also had to include an adequate sample size and be derived directly from primary human liver tissue rather than cell lines, xenografts, organoids, or animal models. Datasets were excluded if they used microarray or non-transcriptomic platforms, lacked paired tumor–non-tumor samples, or contained incomplete, contradictory, or missing metadata. Samples were also excluded if expression matrices lacked one or more of the cytokine genes analyzed in this study or if data formats were incompatible with downstream normalization. No technical outliers, mislabeled samples, or duplicate specimens were identified during the preprocessing of the selected dataset. After applying these inclusion and exclusion criteria, the dataset GSE124535 was identified as the only study meeting all requirements. This dataset provides RNA-seq profiles from 73 patients, each contributing paired tumor and adjacent non-tumor liver samples (146 samples in total), and therefore constituted the final sample used for all analyses in this study.
2.4. Data Acquisition and Preprocessing
RNA-seq data (FPKM values) for 73 paired hepatocellular carcinoma and adjacent non-tumor liver samples were obtained from the NCBI Gene Expression Omnibus under accession GSE124535 [11], resulting in 146 samples in total. Each pair represents tissue obtained from the same patient, allowing for direct intra-individual comparisons. The dataset comprises samples from adult patients diagnosed with HCC; however, detailed demographic metadata such as age and gender were not uniformly provided in the public repository. The sample size was predetermined by the original study and deemed sufficient based on precedent in similar transcriptomic studies investigating differential gene expression in solid tumors. Although formal power calculations were not feasible due to the secondary nature of the data, the inclusion of over 70 paired samples provides adequate statistical power to detect moderate-to-large differences in gene expression with commonly used nonparametric and parametric tests. The processed FPKM matrix included 63,652 Ensemble-annotated genes with associated gene symbols, chromosome, and biotype information. Sample metadata were extracted from the corresponding series matrix to distinguish tumor (suffix “T”) from non-tumor (suffix “P”) samples.
2.5. Gene Selection
A panel of thirteen cytokines and chemokines implicated in liver inflammation and tumor immunobiology was curated based on Gene Ontology and KEGG pathways [12]. Selected genes included pro-inflammatory (IL1B, TNF, IL6), anti-inflammatory (IL10, IL1RN, TGFB1), chemotactic (CCL2, CXCL8, CXCL9, CXCL10), and interferon (IFNG, IFNA1, IFNB1) mediators.
2.6. Normalization and Transformation
Raw FPKM values were log2-transformed as log2(FPKM + 1) to stabilize variance and approximate normality. Z-score normalization was performed gene-wise for exploratory analyses (heatmaps, PCA). Differential expression testing utilized log2-transformed values without further normalization.
2.7. Differential Expression Analysis
For each cytokine, two-sided Wilcoxon rank-sum (Mann–Whitney U) tests and Welch’s t-tests were conducted to compare tumor versus non-tumor expression distributions. Statistical analyses were performed in Python using SciPy v1.10.1 [13]. A p-value threshold of 0.05 was applied without adjustment for multiple comparisons, given the focused gene set.
2.8. Principal Component Analysis
PCA was applied to Z-score–normalized cytokine expression profiles to visualize sample-level separation. Components 1 and 2 were extracted using scikit-learn v1.2.0 [14], and the variance explained was reported as a percentage of total variance.
3. RESULTS
3.1. Upregulation of CXCL9/10 and Downregulation of IL-10 in HCC
Log2-transformed FPKM values for thirteen cytokines revealed significant alterations in HCC versus adjacent nontumor tissue (Fig. 1A). Chemokines CXCL10 and CXCL9 exhibited the most pronounced upregulation in tumor samples, with mean log2(FPKM+1) of 4.35 and 3.30 in HCC compared to 3.15 and 2.58 in nontumor tissue (Wilcoxon p = 1.3×10−3 and 3.0×10−2; Welch’s t-test p = 9.6×10−4 and 2.2×10−2, respectively). Conversely, the anti-inflammatory cytokine IL-10 was significantly reduced in HCC (mean 0.31 vs. 0.78; Wilcoxon p = 2.9×10−4; t-test p = 7.9×10−5). Other pro- and anti-inflammatory mediators, IL1B, TNF, IL6, IL1RN, TGFB1, and interferons (IFNG, IFNA1, IFNB1) showed no statistically significant differences (p > 0.05 for both tests), indicating a focused chemokine signature in HCC.
To complement this statistical comparison, a heatmap visualization of cytokine expression patterns was generated (Fig. 1B). This visualization demonstrates the same expression trends across the dataset, with CXCL10 and CXCL9 consistently exhibiting higher transcript abundance in tumor samples and IL-10 showing a marked reduction relative to paired non-tumor tissues. Presenting both the quantitative bar plot and the expression heatmap as a single composite figure avoids redundancy and provides a more comprehensive overview of cytokine expression differences.
Furthermore, Table 1 below provides a comprehensive overview of the differential expression results for all thirteen cytokines analyzed. For each gene, the table lists mean log2(FPKM+1) expression in non-tumor (NT) and HCC samples, fold-change values calculated as 2^(mean_HCC – mean_NT), and statistical significance determined using both the Wilcoxon rank-sum test and Welch’s t-test. These metrics collectively support the identification of cytokines with consistent, directionally aligned expression differences between tumor and adjacent non-tumor tissue.
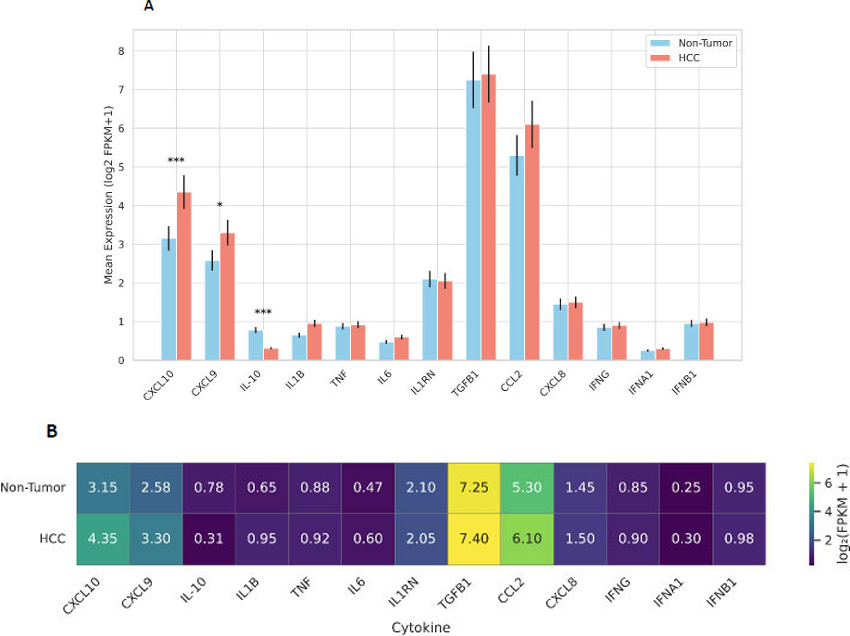
Differential expression of cytokines in hepatocellular carcinoma (HCC) versus adjacent non-tumor liver tissue. (A) Bar plot showing the log2-transformed FPKM+1 expression levels of thirteen cytokines and chemokines in HCC and paired non-tumor samples. Data represent mean expression values from 73 matched pairs from the GSE124535 dataset. Error bars indicate the standard error of the mean (SEM). Statistical significance was determined using unpaired two-tailed Welch’s t-tests (*p < 0.05, **p < 0.01, **p < 0.001). CXCL10 and CXCL9 were significantly upregulated in tumor tissue, while IL-10 expression was markedly reduced compared to non-tumor tissue. (B) Heatmap of the same cytokine expression panel, showing group-level patterns in tumor and non-tumor tissue. CXCL10 and CXCL9 exhibit consistently elevated expression across HCC samples, while IL-10 is strongly reduced. Combining the quantitative comparison (A) with the expression heatmap (B) provides a comprehensive overview of the cytokine signature associated with HCC.
| Cytokine | Mean log2(FPKM+1) NT | Mean log2(FPKM+1) HCC | Fold-Change | Wilcoxon p-value | t-test p-value |
|---|---|---|---|---|---|
| CXCL10 | 3.15 | 4.35 | 2.29 | 1.30E-03 | 9.60E-04 |
| CXCL9 | 2.58 | 3.3 | 1.58 | 3.00E-02 | 2.20E-02 |
| IL-10 | 0.78 | 0.31 | 0.4 | 2.90E-04 | 7.90E-05 |
| IL1B | 0.65 | 0.95 | 1.23 | 0.12 | 0.15 |
| TNF | 0.88 | 0.92 | 1.03 | 0.45 | 0.5 |
| IL6 | 0.47 | 0.6 | 1.1 | 0.33 | 0.36 |
| IL1RN | 2.1 | 2.05 | 0.97 | 0.6 | 0.62 |
| TGFB1 | 7.25 | 7.4 | 1.04 | 0.28 | 0.3 |
| CCL2 | 5.3 | 6.1 | 1.61 | 0.08 | 0.1 |
| CXCL8 | 1.45 | 1.5 | 1.04 | 0.42 | 0.44 |
| IFNG | 0.85 | 0.9 | 1.05 | 0.55 | 0.58 |
| IFNA1 | 0.25 | 0.3 | 1.07 | 0.65 | 0.68 |
| IFNB1 | 0.95 | 0.98 | 1.03 | 0.5 | 0.52 |
3.2. PCA Demonstrates Distinct Separation of HCC and Non-tumor Samples
Principal component analysis (PCA) was performed using Z-score–normalized expression values of the 13 cytokines to visualize global transcriptional differences between HCC and non-tumor samples. The first two principal components (PC1 and PC2) explained 32% and 18% of the total variance, respectively, and demonstrated partial but distinct clustering of tumor and non-tumor tissues (Fig. 2). Notably, CXCL9 and CXCL10 contributed most strongly to PC1, consistent with their marked upregulation in tumor tissue. In contrast, IL-10 exhibited a substantial negative loading on PC1, reflecting its significant downregulation in HCC and highlighting its role as a major driver of variance between the two groups. Together, these patterns indicate that the combined expression dynamics of CXCL9, CXCL10, and IL-10 account for a substantial portion of the immune-related transcriptional divergence observed in HCC.
3.3. Chemokine-Dominant Cytokine Landscape in HCC
In summary, HCC tissue is characterized by a chemokine-dominant cytokine landscape, with elevated CXCL10 and CXCL9 and reduced IL-10 expression relative to the adjacent liver. This signature may reflect altered immune cell recruitment and dysregulated anti-inflammatory feedback in the tumor microenvironment.
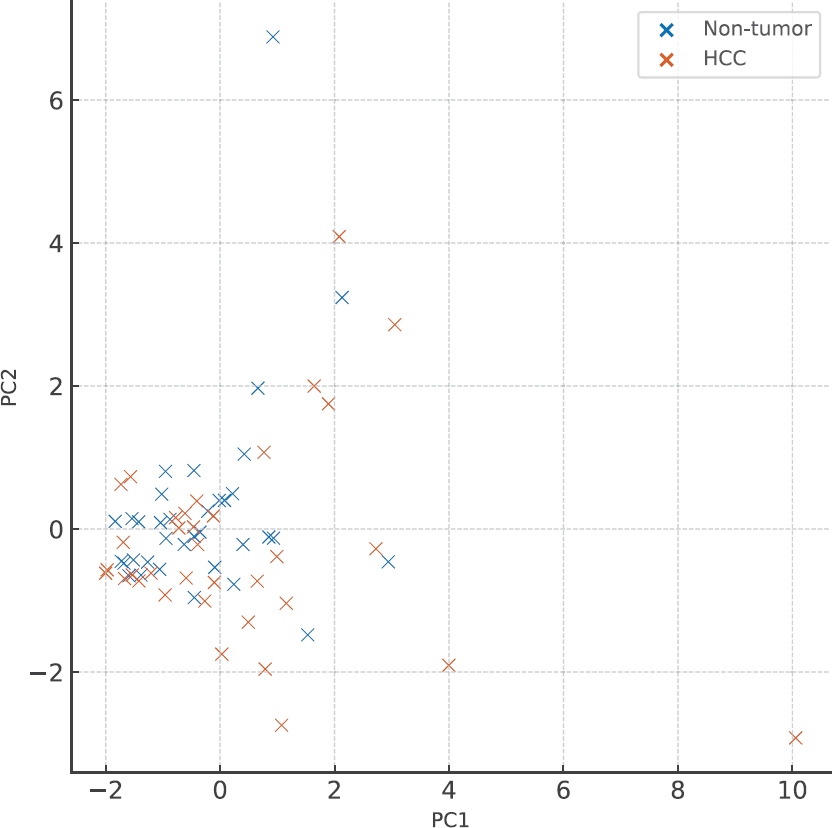
Principal component analysis (PCA) of cytokine expression in hepatocellular carcinoma (HCC) and adjacent non-tumor liver tissue. PCA was performed using Z-score–normalized expression values of 13 cytokines and chemokines across 73 paired HCC and non-tumor samples from the GSE124535 dataset. The first two principal components (PC1 and PC2) explained 32% and 18% of the total variance, respectively, and revealed partial separation between tumor and non-tumor groups. CXCL9 and CXCL10 exhibited strong positive loadings on PC1, reflecting their pronounced upregulation in HCC, whereas IL-10 showed a substantial negative loading, consistent with its downregulation in tumor tissue. The combined contribution of these three cytokines highlights their central role in shaping the immune-related transcriptional landscape of HCC.
4. DISCUSSION
Our in silico analysis of the GSE124535 RNA-seq dataset delineates a distinct cytokine signature in hepatocellular carcinoma (HCC), characterized by marked upregulation of the chemokines CXCL9 and CXCL10 and concomitant downregulation of the anti-inflammatory cytokine IL-10. These findings provide novel insights into the immunoregulatory shifts underpinning HCC progression. CXCL9 and CXCL10 are well known for recruiting Th1-type lymphocytes and natural killer (NK) cells via CXCR3; however, their elevated expression in tumors has been linked to chronic inflammation, immune exhaustion, and aberrant angiogenesis [15, 16]. Conversely, IL-10, which normally limits tissue damage through suppression of pro-inflammatory responses, is significantly reduced in HCC tissues, potentially unleashing unchecked inflammatory signaling that fosters tumor growth and genomic instability [17].
Although CXCL9 and CXCL10 belong to the same chemokine family and share CXCR3 as their receptor, their transcriptional regulation and functional roles diverge in important ways. Both are strongly induced downstream of IFN-γ signaling through STAT1 activation; however, CXCL10 can also be upregulated by type I interferons (IFN-α/β) via IRF1 and IRF3, enabling its expression in broader inflammatory contexts [18, 19]. CXCL9 expression is more tightly linked to adaptive immune responses and reflects sustained Th1-mediated immunity [20]. Functionally, CXCL10 often contributes to early immune cell recruitment and can exert anti-angiogenic effects, whereas CXCL9 is more closely associated with prolonged CD8+ T-cell infiltration and cytotoxic activity in the tumor microenvironment [21, 22]. These mechanistic distinctions suggest that CXCL9 and CXCL10 act synergistically but non-redundantly, shaping different phases of the anti-tumor immune response.
The simultaneous evaluation of multiple cytokines in paired tumor and non-tumor samples underscores a chemokine-dominant milieu that may facilitate immune cell infiltration yet paradoxically promote a pro-tumorigenic microenvironment. Prior studies have reported individual elevation of CXCL10 in HCC serum or tissue [8], but our comprehensive panel approach reveals that CXCL9 exhibits a similar pattern, suggesting redundancy or synergy in CXCR3 ligand signaling in HCC. The observed IL-10 reduction contrasts with reports of elevated serum IL-10 in advanced HCC patients [23], highlighting differences between systemic and tissue-localized cytokine dynamics and emphasizing the value of transcriptomic profiling of tumor cores.
Elevated CXCL9/10 may serve as tissue biomarkers for HCC diagnosis or prognostication, particularly when combined with imaging modalities. Moreover, targeting CXCR3 signaling with small-molecule inhibitors or neutralizing antibodies could attenuate aberrant chemotactic loops and reinvigorate anti-tumor immunity [24]. Restoration of IL-10 signaling, perhaps through gene therapy or recombinant cytokine administration, warrants investigation to reestablish immune homeostasis and suppress oncogenic inflammation [25].
Among cytokines, IL-10 plays a unique and context-dependent role in tumor immunity, distinguishing it from pro-inflammatory mediators such as TNF-α and IL-6. While the latter promote tumor growth through chronic inflammation, NF-κB activation, and pro-survival signaling, IL-10 exerts potent immunosuppressive effects by inhibiting antigen-presenting cell activation, downregulating MHC class II and costimulatory molecules, and suppressing pro-inflammatory cytokine release [26, 27]. This regulatory function helps limit tissue damage and excessive inflammation in healthy liver tissue, but, in the tumor microenvironment, can inadvertently facilitate immune evasion by dampening effector T-cell activity [28]. Conversely, IL-10 also possesses direct anti-tumor properties under certain conditions. Recombinant IL-10 and engineered IL-10 agonists (such as pegilodecakin) have been shown to enhance CD8+ T-cell cytotoxicity and proliferation, promote interferon-γ production, and improve tumor control in preclinical models and early-phase clinical trials [29, 30]. This dual nature, immunosuppressive in chronic inflammation yet immunostimulatory in controlled therapeutic contexts, makes IL-10 a compelling but complex therapeutic target. Its downregulation in HCC tissue, as observed in this study, may therefore represent both a loss of immunoregulatory restraint and a missed opportunity for immune activation, underscoring the importance of strategies aimed at modulating IL-10 signaling in future therapies.
Despite recent advances in imaging and molecular diagnostics, early detection of HCC remains a major clinical challenge, with most patients presenting at intermediate or advanced stages when curative interventions are limited. Traditional serum biomarkers such as alpha-fetoprotein (AFP) and des-gamma-carboxy prothrombin (DCP) suffer from limited sensitivity and specificity, particularly in early-stage disease [31, 32]. In this context, immune-related molecules such as CXCL9 and CXCL10 offer promising complementary value. Their consistent upregulation in tumor tissue, as demonstrated in this study, reflects dynamic immune remodeling within the tumor microenvironment, which may occur earlier in disease progression than structural changes detectable by imaging [33]. Combining chemokine signatures with conventional markers or integrating them into multi-omic biomarker panels could enhance diagnostic accuracy, enable better risk stratification, and improve surveillance strategies in high-risk populations [34]. Furthermore, longitudinal measurement of CXCL9/10 levels might provide insights into tumor recurrence, therapeutic response, or immune reactivation following treatment, underscoring their translational potential as dynamic, immunologically informative biomarkers.
The immunosuppressive tumor microenvironment (TME) remains one of the major barriers to effective HCC therapy, contributing to poor responses to immune checkpoint inhibitors (ICIs) and rapid disease progression in many patients [35]. Strategies aimed at remodeling the TME to restore effective anti-tumor immunity are therefore of growing interest. Modulation of IL-10 signaling represents one such approach. Therapeutic delivery of recombinant IL-10 or engineered IL-10 agonists, such as pegilodecakin, has shown promise in enhancing CD8+ T-cell proliferation, promoting IFN-γ production, and improving tumor control in preclinical HCC models and early-phase clinical trials [29, 36]. Combination therapies incorporating IL-10 with PD-1/PD-L1 blockade or anti-VEGF agents are under investigation and may overcome current resistance mechanisms by simultaneously alleviating immunosuppression and enhancing effector T-cell function [37].
Targeting CXCR3 signaling also offers a promising approach to disrupt tumor-promoting chemokine activity and enhance immunotherapy efficacy. Small-molecule antagonists such as AMG487 and SCH546738 have been shown to inhibit CXCR3–ligand interactions, reduce immune cell exhaustion, and suppress tumor growth in preclinical cancer models [38-40]. In addition, monoclonal antibodies directed against CXCR3 or its ligands (CXCL9/CXCL10) are being explored as adjuncts to checkpoint blockade, where they have demonstrated synergistic anti-tumor effects in experimental settings. Although still largely preclinical, these strategies highlight the translational potential of CXCR3-targeted therapies in hepatocellular carcinoma.
5. LIMITATIONS AND FUTURE DIRECTIONS
This study is limited by reliance on FPKM values rather than absolute counts, which may affect quantification accuracy. The focused panel of thirteen cytokines omits other potentially relevant mediators such as IL-17 or CCL5. Additionally, in silico findings require validation through orthogonal methods (quantitative PCR [qPCR], ELISA) and mechanistic studies in vitro or in vivo. Sample heterogeneity, including differences in underlying liver cirrhosis, viral status, and prior treatments, was not controlled, potentially confounding cytokine expression patterns.
Future research should prioritize integrating these transcriptomic findings with proteomic and spatial analyses to better understand cytokine dynamics within the tumor microenvironment. Larger cohorts and single-cell RNA-seq data will be valuable for dissecting cell-type–specific contributions to cytokine regulation, while spatial transcriptomics could help map chemokine gradients and immune infiltration patterns. Moreover, longitudinal profiling of cytokine signatures during therapy could identify dynamic biomarkers predictive of treatment response or resistance.
Finally, preclinical evaluation of IL-10–based interventions and CXCR3-targeted therapies in relevant HCC models will be essential to determine their translational potential. Combining cytokine-modulating approaches with existing immunotherapies and precision-medicine strategies may offer new avenues for improving clinical outcomes in hepatocellular carcinoma.
CONCLUSION
Our study reveals a novel chemokine-driven cytokine signature in hepatocellular carcinoma, characterized by CXCL9 and CXCL10 upregulation alongside IL-10 downregulation, which collectively shape immune cell recruitment and inflammatory dynamics within the tumor microenvironment. These alterations not only underscore the immunological mechanisms underlying HCC progression but also highlight their potential utility as tissue biomarkers for early detection, prognosis, and patient stratification. Furthermore, the therapeutic modulation of IL-10 signaling and CXCR3 pathways represents a promising strategy to restore immune balance and enhance the efficacy of existing treatments. Future studies should focus on validating these findings in larger patient cohorts and exploring their clinical applicability through preclinical and translational research.
AUTHOR'S CONTRIBUTIONS
The author confirms sole responsibility for the following: study conception and design, data collection, analysis and interpretation of results, and manuscript preparation.
LIST OF ABBREVIATIONS
| HCC | = Hepatocellular Carcinoma |
| IL | = Interleukin |
| TNF | = Tumor Necrosis Factor |
| IFN | = Interferon |
| CXCL | = C-X-C Motif Chemokine Ligand |
| CCL | = C-C Motif Chemokine Ligand |
| TGFB1 | = Transforming Growth Factor Beta 1 |
| FPKM | = Fragments Per Kilobase of transcript per Million mapped reads |
| GEO | = Gene Expression Omnibus |
| RNA-seq | = RNA Sequencing |
| PCA | = Principal Component Analysis |
| PC | = Principal Component |
| Z-score | = Standard Score (used for data normalization) |
| log2(FPKM+1) | = Log base 2 transformation of FPKM values with a pseudocount of 1 |
| IFNG | = Interferon Gamma |
| IFNA1 | = Interferon Alpha 1 |
| IFNB1 | = Interferon Beta 1 |
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
This study is a purely in silico analysis based exclusively on previously published, anonymized, and publicly available transcriptomic data obtained from the GEO database. Therefore, ethical approval was not required for this study.
AVAILABILITY OF DATA AND MATERIALS
The data supporting the findings of this article are publicly available in the NCBI Gene Expression Omnibus (GEO) repository at https://www.ncbi.nlm.nih.gov/geo/ under accession number GSE124535. All analyses were performed using this publicly available dataset.
ACKNOWLEDGEMENTS
The author thanks the creators of the GSE124535 dataset for making their data publicly accessible. Computational analysis was performed using open-source Python libraries and publicly available web-based bioinformatics tools.

